

今村祐嗣教授 研究のあゆみ
2010年3月の最終講義より
まえがき
京都大学教授今村祐嗣先生は、誠に名残惜しいことではございますが、来る平成22 年3 月
31 日をもちまして、定年退職されることになりました。
本事業会では、先生のご退職にあたり、先生へのお礼および感謝の意をこめて、最終講義
を企画いたしました。最終講義では、「木を観て、木に探す -私の木質科学研究-」という
演題で、先生のこれまでのご研究に関するお話を頂く予定です。
本冊子では、先生のご略歴、研究の歩み、先生が主査となって取得された学位論文の概要
および卒業生からのお礼・感謝の言葉を取りまとめました。ご参加の皆様におかれましては、
ご一読いただき先生のこれまで幅広いご業績ご活躍を振り返っていただければ幸いでござい
ます。
最後に、先生の輝かしいご業績を讃えるとともに、長年にわたるご指導に深謝し、先生の
今後のさらなるご発展と益々のご健勝を心よりお祈り申し上げます。
平成22 年3 月吉日
今村祐嗣先生定年退職記念事業会
目次
3 1971 - 1974 年
[ピットはどのように形成されるのか]
1974 - 1982 年
[スギ天然シボの組織構造とシボ形成への考察]
[過熟材]
[曲げ木]
1982 - 1991 年
[腐朽材のフラクトグラフィー]
[木質ボードの腐朽と虫害]
[木材の化学修飾]
[ピットと水分移動]
[注入性向上技術]
[劣化診断]
1991 - 2000 年
[低分子フェノール樹脂処理]
[横圧縮前処理法]
[リモートセンシングでシロアリの行動を探る]
[木材の風化]
[木材の屋外保護塗料]
[木材劣化の気候指標]
2000 年以降~
[シロアリを匂いで検出]
[木炭のナノ構造]
卒業生からのお礼・感謝の言葉
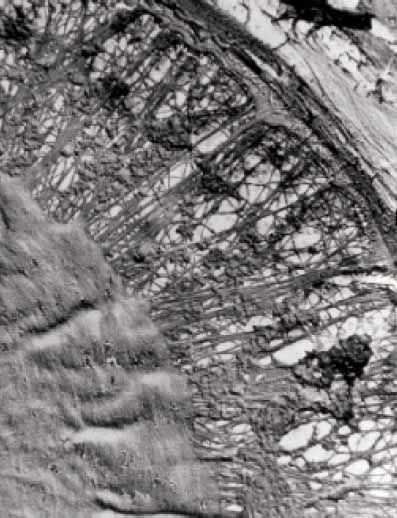
京都大学農学研究科林産工学専攻の博士過程で、故 原田 浩教授(木材構造学研究室)の指導の下で、電 子顕微鏡による木材細胞壁の形成過程、とくに樹液や水分通導の役割を担っている壁孔(pit)の形成と微細 構造の研究を行った。電子顕微鏡の観察手段であるダイレクトレプリカ法の改良法を開発して、分化過程に おける木材の細胞壁でのセルロースミクロフィブリルの堆積のようすから壁構造の解明に取り組んだ。この 分化過程の細胞壁観察法は、その後も細胞壁の微細構造観察の有力な研究手法として用いられた。
“Studies on the Development and Ultrastructure of Pits in Coniferous Xylem”で、1975 年1 月に京都大学農学博 士の学位を授与される。この研究は、針葉樹仮道管の壁孔の形成と微細構造を分化中の細胞壁内表面の電子 顕微鏡的観察から明らかにしたもので、分化中細胞の原形質分離と凍結乾燥法を試料調整法に、ダイレクト レプリカ法を観察手法として行った。特に明らかにした点は、①トールス、マルゴという壁孔膜のミクロフィ ブリル構造がすでに一次壁の段階で形成されていること、その形状のまま分化段階では非晶物質で充填され ているが、分化終了時に開孔されること、②内表面側に形成される壁孔縁を順次観察したところ、S1 層の 形成段階でミクロフィブリルが一定間隔で同一方向に並んだミクロラメラが連続的に堆積し、それらが孔口 で迂回することから壁孔縁が形成され、その後のS2 層でのミクロラメラの堆積で成長すること、③放射柔 細胞に形成される分野壁孔の細胞壁構造についても観察を行い、ランダム配向の壁層にはさまれて明瞭な交 差構造をしたミクロフィブリルの層が存在すること、等である。
[ピットはどのように形成されるのか]
細胞が伸長生長していく過程でつくられる一次壁の形成が終わると、細胞の形は定まり続いて二次壁 の堆積が始まる。一次壁の形成が終了した段階で将来ピットになる領域はすでに円形に仕切られてお り、しかも中央のトールスと周辺のマルゴの区分ができている。さらにマルゴには放射状に配列したミ クロフィブリルの存在さえうかがうことができる。マルゴの部分は非セルロース様物質で密に充填され ているが、これを化学的に除去してみると、完成されたメンブレンとまったく同等の仕組みをすでに認 めることができる。
マルゴにおけるミクロフィブリル束の放射状配列については、本来網目状配列として形成されたもの がトールスの移動によって引っ張られ、二次的に再配列したものではないかという考え方もあった。し かし、すでに分化段階で形態形成がなされていることが明らかになったことで、これは細胞の壁形成機 能によってもたらされたものであることは明らかである。このことは、仮道管と放射組織とが交差する 分野領域においても、仮道管側に放射状に伸びるミクロフィブリルが観察されることからも示されてい る。
水分移動の制御に機能的なトールスとマルゴの形が、細胞壁の形成段階のきわめて初期につくられて いるのは、樹木の細胞壁形成の不思議な一項であろう。
分化段階の間を通して非晶物質で充填されていたマルゴは、二次壁の最内層の形成が終了すると分解 され、メンブレン構造ができあがる。この現象は、広葉樹の道管で分化期間中は充填されていた穿孔板 が開孔するのと同様なオートリシスの一つと考えられる。
 |
 |
充填されたピットメンブレン |
開孔途上のピットメンブレン |
当時の木材構造学研究室のスタッフは、原田教授以下、佐伯 浩助教授(後 教授、現 京都大学名誉教授)、谷口助手(後 新潟大学教授、現 同名誉教授)、野渕 正助手(後 教授、現 京都大学名誉教授)であった。
1974 - 1982 年
昭和49 年(1974 年)4 月から施設拡充された奈良県林業試験場(工藤俊夫所長 現 奈良県森林技術センター)
木材化学課に勤務し、木材、特に人工林率がきわめて高い奈良県の地域材の材質・強度の性能評価と利用開
発の研究に従事した(大学時代の電子顕微鏡によるミクロン、オングストロームの世界から、吉野の山で立
木を伐採して自ら帯鋸で製材し、100 トン容量の強度試験機で実験するという大きな転換であった)。
木材材質の研究では奈良県産のスギ材、特に間伐材の強度・材質を保育方法(密植多間伐の吉野方式と比
較的粗植の十津川流域の方式)と関連させて解析した(小野広治、山口和道、海本 一氏らと)。また、集成
材の構造材利用の発展期にあたっていたこともあり、集成材の強度性能の解析、特に大断面集成材のせん断
破壊の解析等の研究業務に参画した(中西祺周氏らと)。
一方、京都大学木材研究所の島地 謙教授(京都大学名誉教授、故人)が開発した年輪内マーカー法とし
てのピン打ち法を多くの樹種に展開する研究を共同で行うとともに、傷害組織形成を樹木の傷害によるレス
ポンスとしてとらえて特異な細胞構造の観察を行った。また、スギ変種である天然シボ材(丸太表面が凸凹
にしゅう曲するなど品種ごとに独特の形状をもち、遺伝的にそれらの形質をひきつぐもの)の組織的特徴の
解明に着手し、放射組織形態の特徴的分類から品種が識別できる手法を開発した(岡崎 旦氏らと)。
[スギ天然シボの組織構造とシボ形成への考察]
スギ天然シボ
スギは老齢になれば年輪がしゅう曲し、樹幹表面にシワを現す性質があるが、若齢であっても年輪画
しゅう曲し幹の表面に凸凹状の、いわゆる“シボ”をつくるものがある。これが天然シボである。この
天然シボは、スギ以外にヒノキやケヤキなどにも発生し、以前から磨丸太にして床柱に利用されてきた。
スギ天然シボ丸太を建築物に使用した例は古く、京都の大徳寺黄梅院の茶室「昨夢軒」の床柱のように
約400 年前にさかのぼることができる。
スギ天然シボは、年輪がへこむように落ち込み、幹の表面に溝状のしゅう曲が縦方向に形成される様
相を呈するのが入シボ、逆に隆起したシボ模様を示すのが出シボと呼ばれる。入シボは立地条件など環
境要的因子によって誘発されることが多く、出シボは先天的な遺伝形質として次代に伝わる特徴をもつ
とされる。
天然シボのスギは品種ごとに幹表面のシボ模様が異なり、チリメン状とかコブ状とかの独特の凸凹を
もっている。天然シボの品種は、発見者や発見場所からそれぞれ独自の名称がつけられているが、幹表
面にシボ模様によって大別すると、細かいチリメン状のシボをもつヒロガワラやウメダ、大きなコブ状
のウチアイやヨシベ、両者の混ざったナカゲンやウンガイに分類される。
天然シボスギの組織の特徴-放射組織と品種
一般的の天然しぼのスギでは、放射組織の接線断面での形状が一般のスギに比べてより丸くかつ大き
く、また、複列の放射組織あるいは不規則な形をしたものが観察された。もちろん、材中に占める放射
組織の体積割合も増大している。これらの放射組織の特徴は、老齢木あるいは外的な傷害や病虫害を受
けた材部に見られる特徴と共通している。
天然シボの組織構造における放射組織の特徴で興味深いのは、品種あるいは幹表面でのシボ模様にそ
れらの特徴が対応していることである。すなわち、複列あるいは多列の放射組織が不規則に集結して現
れるタイプは幹表面に細かいチリメン状のシボをもつヒロがワラやウメダに、複列の放射組織が分散し
ているタイプは細かいシボとコブが混在しているナカゲンに、複列あるいは多列の細胞が部分的に集
まって出現するタイプはナカゲンより発達したチリメンシボをもつクロにそれぞれ対応して観察され
る。
特筆すべきことは、これらの放射組織の特徴は品種固有であるとともに、幹表面において年輪がまだ
しゅう曲し始めていない段階でも観察されることである。すなわち若齢木の段階で将来のシボ形状を予
測する因子として利用できるといえる。
シボ形成のメカニズム
通常の木部組織においては、幹の肥大生長に合わせて、形成層の紡錘形始原細胞が半径方向だけでな
く適宜接線方向に分裂し、円周の拡大に対応するよう自己調節を行っている。すなわち、紡錘形始原細
胞は偽横分裂によって接線方向に数を増やすが、樹木に備わっている自己調節機能がはたらいて半径方
向の拡大に応じたバランスを保っている。こういった偽横分裂した紡錘形始原細胞が機能するかどうか
は、放射組織始原細胞との接触頻度によって左右されているらしい。
天然シボ品種の場合、遺伝的指令によって放射組織始原細胞が過剰に誕生し、その結果、偽横分裂し
た紡錘形始原細胞が数多く誕生し、接線方向の細胞分裂が半径方向のそれとのバランス以上に機能し
て、シボが生じたのではないだろうか。
 |
 |
| ナカゲンの天シボ丸太 | ウメダの接線断面 |
 |
 |
| クロの接線断面 | 通常のスギ材の接線断面 |
さらに、老齢に達した樹木の材質(過熟材とも称する)に関心を抱き、老齢スギの銘木である春日杉(奈
良春日奥山)の解剖学と材質解析を行った。
[過熟材]
老齢化した樹木の材部はどういう性質を備えているのだろうか。一番の特徴は年輪幅がきわめて狭く
なることである。大体250 年以上になると年輪幅が0.5 ミリ前後の超過密の年輪を構成するようになる。
これぐらいの年輪になると木口面で数えると1 年輪に10 細胞くらいしか並んでいない。ちなみにスギ
の成熟した材の平均年輪幅は3 ~ 4 ミリ前後が多いが、この場合は100 細胞以上を数えることができる。
すなわち、細胞の数では成熟材の10 分の1 くらいしか1 年に生長していないことになる。
顕微鏡で細胞の形をみてみると、早材の細胞は直径が大きくかつ細胞の壁がきわめて薄いが、一方で
晩材では径が小さく細胞壁は極端に厚い。すなわち、早材と晩材のコントラストが著しい。老齢化した、
いわゆる過熟材の早材部分は強度も密度も低いが、この理由は未成熟材と同様に木材を構成している細
胞壁のフィブリル傾角が緩やかになっていることと、成分的にセルロース量が少なくてリグニンが多い
ことに起因している。
このスギの老齢化した部分では、それまで真円状に生長してきた年輪が波打つようになってくる。す
なわち年輪がしゅう曲する。老齢化して年輪が波打つ現象はレッドウッドやベイスギなどにもみられる
が、スギの場合がとくに顕著であり、板目面には笹杢など美しい木目をつくり出す。国産材とくにスギ
の利用開発が重要なことはいうまでもないが、難しい、困ったとシワを寄せさせてはならない。
 |
 |
| 春日スギの木口切片の軟X線(左) | 板目材の笹杢(右) |
山中勝次氏(奈良県林業試験場からホクト産業㈱、現 京都菌類研究所長)とは、SEM を用いた硅化木の
組織構造の研究を行い、この成果は後の無機質複合化木材の組織構造面からの裏付け研究に関連してくるこ
とになる。
またこの時期、京都大学木材研究所の則元 京氏(現 京都大学名誉教授)の研究グループの一員として、
曲げ木の組織構造的解明に取り組んだ(和田 博、飯田生穂、林 昭三氏らと)が、これ以降、木材組織学の
立場から物性研究者らとの共同研究を行うことになる。
[曲げ木]
マイクロ波の利用というと、すぐ思い出すのは電子レンジで、この場合は中にいれた物が周囲から発
信された電波を吸収して振動し、その時の摩擦で電波のエネルギーが熱に変換される仕組みになって
いる。特に誘電率の高い水(一般の物質の中でも誘電率が高いとされる陶磁器に比べても約10 倍)は、
電波を吸収して温度が上がりやすい傾向をもっている。
かなり昔のことになるが、当時同じ研究仲間であった則元京 京大名誉教授(現、同志社大学教授)
らとマイクロ波加熱を利用した曲げ木を研究したことがある。水を含んだ木材を電子レンジの中で加熱
して乾燥と同時に曲げ加工するもので、従来の方法に比べてより大きな変形が可能になった。写真はス
ギ材を加熱して型に沿ってかなり小さな曲率にまで曲げ加工し、乾燥固定後に湾曲内側の圧縮部分を観
察したものである。木材の細胞壁自体が折れ曲がるように波状にしゅう曲し、しかも変形は圧縮破壊と
異なり細胞壁に均等に分散していた。このしゅう曲が圧縮破壊と異なるのは、再度水分を吸収させると
曲げ加工した材は直材に戻り、細胞壁の“しわ”もほとんど消失したことで証明されている。すなわち、
マイクロ波加熱によって木材細胞壁のマトリックス成分が軟化し、変形が容易に生じたものと解釈され
た。擬音語表現の得意な則元教授が、“木がぐにゃぐにゃになった”と当時表現されていたことを憶え
ている。
 |
 |
| 曲げ木(左) | 圧縮部におけるスギ細胞壁のしゅう曲(右) |
また、布村昭夫氏(北海道立林産試験場、北海道東海大学教授、現 退官)と一緒に行ったインサイジン
グした製材の強度性能の研究は、その後の木材保存研究の嚆矢ともいえるものであった。
この間、昭和51 年から企画情報係を併任して、研究報告の定期発行など試験場の研究基盤の強化に従事
した。
1982 - 1991 年
昭和57 年(1982 年)2 月に京都大学木材研究所附属木材防腐防虫実験施設(西本孝一教授、高橋旨象助教授、
角田邦夫助手;当時)の助手に、時限性部門の補強要員として採用される。同施設はその後改組されて高耐
久性木材開発部門(西本孝一教授、角谷和男教授、その後高橋旨象教授)になったが、1988 年9 月に同部
門の助教授に昇任した。この間の研究の主たる目標は、防腐防虫研究の材料、建築への展開ということに要
約できる。防腐防虫実験施設および新設された高耐久性研究部門における研究内容を以下にまとめる。この
うち、いくつかについては、その後の複合材料分野での仕事に結び付いた。
●高橋旨象教授の開発した『腐朽菌作用下での曲げクリープ試験法』を発展させ、木材および木質材料の
腐朽による強度性能の変化を体系化した。これに関連して、腐朽材のフラクトグラフィー、すなわち破
壊面の特徴から腐朽のタイプや程度を明らかにする研究法を開発した。
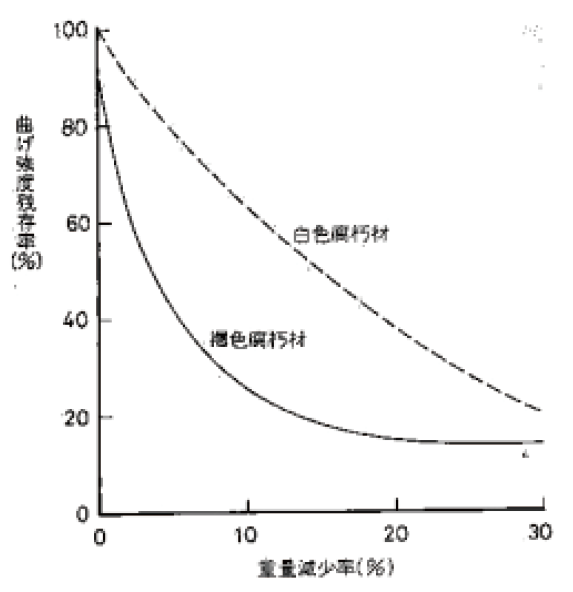 |
 |
| 腐朽材(素材)の曲げ強さ | 強制腐朽の様子 |
[腐朽材のフラクトグラフィー]
木材腐朽菌は褐色腐朽菌と白色腐朽菌に大別される。このうち褐色腐朽菌は針葉樹材を特に劣化さ
せ、セルロースを選択的に分解し大きな強度低下を引き起こすことから建築物等の主要な劣化微生物で
ある。白色腐朽菌はセルロースとリグニンとを同時に分解するため強度低下に与える影響は褐色腐朽菌
よりも小さいが、広葉樹材をより好み、樹木の腐れなどでは一般的に認められる菌類である。
褐色腐朽菌と白色腐朽菌の木材成分の分解様式は、腐朽した木材の組織構造にも顕著に認められ、破
断面の特徴を表している。
●木質ボードの腐朽劣化の機構について検討を行い、耐久性を向上させる要因を解析する。この中で、腐
朽菌によるボードの接着層への侵入機構をはじめて明らかにし、接着剤の種類によってその程度が大き
く異なることを示した。さらに、この研究の延長として、木質材料部門グループと接着剤中への薬剤混
入法の開発に取り組み、高耐久性発現のためのシステムつくりを行った。この手法は、その後、難燃木
質材料の開発へと展開することになる。
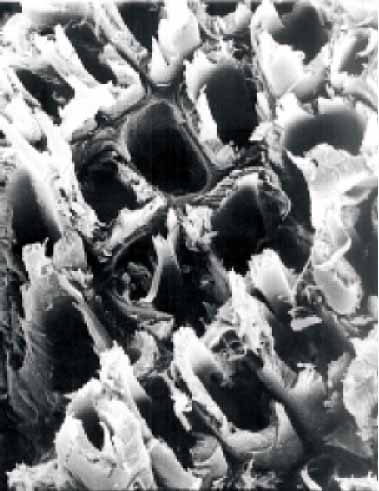 |
 |
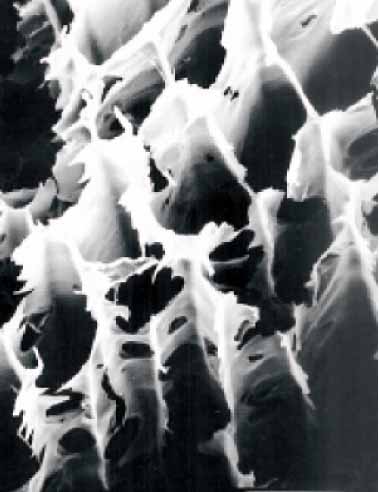 |
| 健全材 | 褐色腐朽 | 白色腐朽 |